15 studi forestali del 2025, scelti e raccontati da SISEF – Parte 5

Questo post fa parte della Rubrica “15 studi forestali del 2025, scelti e raccontati da SISEF – Parte 3”, se ti sei perso i primi episodi puoi recuperarli qui:
Cosa vuol dire, concretamente, gestire un bosco in modo sostenibile? Nei documenti di pianificazione lo leggiamo – o lo scriviamo – spesso, ma poi arriva il momento difficile: tradurlo in numeri, in scelte operative, in ettari da destinare a produzione o a tutela. I tre studi che raccontiamo in quest’ultimo post dedicato alle ricerche forestali più interessanti del 2025 provano a chiudere proprio quel divario. Il primo parte una teoria elegante dell’ecologia, il principio di equivalenza energetica, e la trasforma in un “righello” per leggere la struttura di un bosco e definire obiettivi di intervento. Il secondo mette insieme decenni di sperimentazioni e si chiede se il diradamento renda davvero i boschi più resilienti a siccità, insetti, vento e fuoco. Il terzo alza lo sguardo sulla pianificazione: quali combinazioni di gestione forestale intensiva, estensiva e assenza di gestione massimizzano la biodiversità, e per quali gruppi tassonomici?
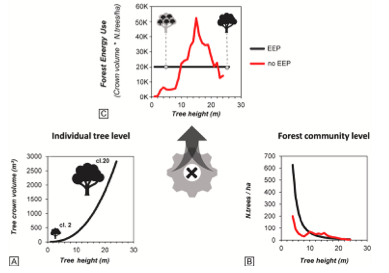
1) Operationalizing closer-to-nature forest management: application of the energy-equivalence principle in a holm oak (Quercus ilex L.) Mediterranean forest (Pasqualotto et al., Forest Ecology and Management)
Un team di ricerca dell’Università di Padova, dell’Agenzia Regionale Fo.Re.S.T.A.S. e dello studio forestale D.R.E.AM. Italia ha lavorato nei lecceti del Supramonte (Sardegna), partendo da un’idea semplice: se un bosco riesce a mantenere nel tempo sia molte piantine sia alcuni grandi alberi, significa che le risorse disponibili sul sito (luce, acqua, nutrienti) vengono “spese” in modo equilibrato tra individui piccoli, numerosissimi ma poco esigenti, e individui grandi, pochi ma molto costosi in termini di energia e risorse. Il principio di equivalenza energetica (EEP) formalizza questa idea: all’aumentare della dimensione, ogni albero richiede più risorse, quindi per restare in equilibrio la foresta può “permettersene” meno, producendo una relazione prevedibile tra abbondanza e dimensione media delle piante.
Per applicare questo principio a ogni foresta, però, serve un passaggio tecnico cruciale: bisogna stimare come cresce il consumo di risorse con l’aumentare delle dimensioni degli alberi. Gli autori lo fanno misurando altezza, diametri e dimensioni della chioma nella lecceta di Montes e usando il volume di chioma come indicatore della superficie fogliare, e quindi della “spesa energetica”, di ogni pianta. Da queste misure ricavano una relazione di potenza molto robusta tra volume di chioma e altezza: l’esponente di scala è circa 3, con un R² intorno a 0,89. Applicando questa power law è possibile calcolare la pendenza attesa della distribuzione delle dimensioni in condizione di equivalenza energetica, cioè il numero che descrive quanto rapidamente diminuisce il numero di alberi quando aumenta la loro dimensione (β = −2,44).
A questo punto entra in scena il confronto con la foresta reale, misurata con 70 rilievi e transetti dedicati alla rinnovazione. Nella parte vetusta della lecceta, la distribuzione delle dimensioni degli è vicina alla previsione EEP, ma non coincide: la pendenza stimata è β = −1,99 (R² = 0,95). Nelle isole di senescenza inserite all’interno della foresta regolamente gestita, lo scarto è molto più marcato: β = −1,36 (R² = 0,89). Gli autori traducono lo scarto tra “foresta reale” e “foresta EEP” in indicatore quantitativo, una traccia matematica esperesso in termini di numero di alberi per ciascuna classe dimensionale che un tecnico può conoscere e, se vuole, adottare. Nel bosco vetusto analizzato qui, la categoria “rinnovazione” (diametro < 10 cm) dovrebbe aumentare del 75% rispetto allo stato attuale; anche le classi maggiori dovrebbero crescere (spessina +39%, perticaia +14%), mentre le classi grandi e molto grandi dovrebbero ridursi di circa il 30% ciascuna per riallocare risorse verso le dimensioni più piccole. Nelle isole di senescenza il quadro è ancora più netto: rispetto alla struttura EEP, la rinnovazione manca quasi dell’87%, mentre la classe di perticaia risulta in eccesso (+54%).
Qui la teoria diventa diagnosi: non solo c’è poca rinnovazione, ma manca esattamente quella parte di popolamento che dovrebbe sostenere la transizione verso una struttura più complessa. Gli autori collegano la carenza di individui giovani soprattutto al disturbo da erbivori, perché in aree recintate, dove il pascolamento è escluso, osservano uno strato più denso di giovani piante. Da questo punto di vista, specialmente in area mediterranea, gli arbusti possono creare microclimi più miti e proteggere le plantule forestali dagli erbivori; nella lecceta, ginepro rosmarino ed erica che occupano il 30–60% dello strato di rinnovazione con diametro > 1 cm in molte radure, e la densità della rinnovazione di leccio è proporzionale alla presenza di arbusti.
2) Potential of thinning to increase forest resilience and resistance to drought, pest, windstorm and fire: A meta-analysis (Chagnon et al., Forest Ecology and Management)
Il secondo studio risponde una domanda che in molti boschi europei è diventata pressante: il diradamento funziona sempre come misura di adattamento climatico, o la sua efficacia dipende dal contesto dire? Questa meta-analisi riassume i risultati di 50 studi e 246 casi, prendend in esame sia la capacità della foresta di resistere agli impatti (resistenza) che di recuperare dopo l’impatto (recovery) o ritornare ai livelli di performance precedenti (resilienza). Al di là degli indicatori specifici per ciascun disturbo (ad esempio i cambiamenti di ampiezza negli anelli di accrescimwento annuali per lo stress da siccità), In tutti i casi, l’effetto del diradamento è sempre stato sintetizzato come standardized mean difference tra trattamento e controllo non diradato.
Il risultato medio, aggregato, è positivo: l’effetto complessivo del diradamento sulla resilienza è stimato pari a 1,08 (intervallo di confidenza al 95%: 0,45–1,70). Detto in modo semplice, nella media dei casi considerati, i popolamenti diradati mostrano una risposta più favorevole rispetto ai controlli non diradati, ma con una variabilità di comportamenti piuttosto ampia.
Quando si separano i disturbi, emergono differenze importanti. Per la siccità, l’effetto medio è moderato ma significativo, e simile sia per resistenza che per recupero. Ridurre competizione e densità aiuta gli alberi durante lo stress idrico e favorisce la ripresa della vitalità dopo l’evento siccitoso. Per insetti e patogeni, la storia è più complessa: l’effetto medio complessivo è superiore, ma con un intervallo che comprende anche lo zero. Qui la resistenza cresce in modo più netto, mentre l’effetto sul recupero è più piccolo e non sempre significativo. L’interpretazione è che il diradamento può ridurre la suscettibilità o la gravità dell’attacco (magari perché migliora vigore, microclima, o riduce la continuità di chioma), ma una volta che l’attacco è avvenuto, non sempre accelera il ritorno allo stato precedente.
Il vento è il disturbo per cui i tecnici non si aspetterebero un “sì o no” netto, perché l’ipotesi è ambivalente: meno alberi significa meno effetto vela complessivo, ma anche maggiore esposizione per gli individui rimasti e, talvolta, chiome più sviluppate. In media, qui la meta-analisi trova un effetto praticamente nullo. Anche con la distinzione resistenza/recupero, l’effetto resta vicino allo zero. Questo non significa che il diradamento non sia mai efficace, ma solo che, aggregando studi e contesti, non emerge un vantaggio generale e robusto del diradamento contro il rischio da vento. Infine il fuoco: qui l’effetto medio stimato è grande, ma l’incertezza è enorme. La ragione risiede più che altro nelle dimensione del campione: nella banca dati sono pochissimi i casi disponibili per questo disturbo, quindi l’intervallo dice più “non abbiamo abbastanza evidenze” che non “l’effetto è estremamente variabile”.
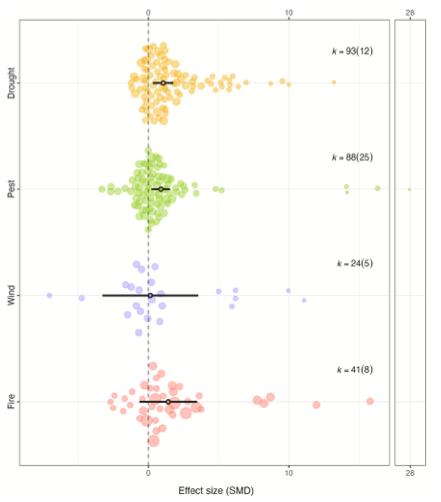
Nelle conclusioni ,gli autori propongono una lettura operativa: nelle foreste temperate e boreali di Nord America ed Europa, il diradamento, nel complesso, tende ad aumentare resistenza e resilienza a più stressor, e soprattutto non emergono effetti significativamente negativi della sua applicazione sugli indicatori di resistenza e recupero post-disturbo. Almeno con rimozioni di area basimetrica relativamente basse (≤30%), il trattamento può essere considerato a basso rischio anche rispetto allo schianto da vento, aprendo a interventi mirati dove siccità, insetti e in alcuni contesti anche fuoco stanno crescendo. Allo stesso tempo, il diradamento da solo non basta e spesso va integrato con altre strategie (per esempio con trattamenti fitosanitari sulle delle ceppaie in presenza di Armillaria, o con fuoco prescritto e gestione dei combustibili in contesti di elevato pericolo di incendio). Inoltre, i benefici osservati a scala di popolamento hanno una durata limitata, quindi vanno letti come una finestra temporale utile per impostare strategie adattative di più lungo periodo.

3) Sustainable forest planning: Assessing biodiversity effects of Triad zoning based on empirical data and virtual landscapes (Duflot et al., PNAS)
Il metodo di zonizzazione Triad, sperimentato in USA e canada, si basa su un’idea semplice ma potente: in una parte di territorio la gestione forestale è ad alta vocazione produttiva (gestione intensiva), in un’altra parte la gestione è estensiva (spesso attraverso selvicoltura naturalistica, continuous cover forestry, o attenta a servizi ecosistemici di regolazione), e nella terza il bosco è lasciato alla dinamica naturale (non gestito). Il presupposto implicito è che la gestione estensiva o integrata sia il migliore compromesso: meno impatto, più biodiversità, e produzione comunque garantita legno. Rémi Duflot e i suoi colleghi, tra cui Giovanni Trentanovi del CNR-IRET e Sabina Burrascano dell’Università Sapienza di Roma, mettono questo presupposto alla prova su faggete eutrofiche submontane europee, usando dati empirici con campionamento multi-taxa e un esperimento “virtuale” costruito per simulare paesaggi con diverse percentuali di ciascuna categoria, con 66 combinazioni possibili e 5000 ricampionamenti di 20 plot per combinazione, il tutto su nove regioni in quattro Paesi (Repubblica Ceca, Francia, Germania e Italia) e su sei gruppi tassonomici: uccelli, coleotteri saproxilici, piante vascolari, briofite epifite, licheni epifiti e funghi lignicoli.
Le risposte della biodiversità alle diverse combinazioni del metodo Triad vengono stimate con GAM bidimensionali lungo i gradienti di composizione. Il risultato più chiaro è anche quello più controintuitivo: la diversità multi-tassonomica a scala di paesaggio (gamma) raggiunge il massimo in un paesaggio composto da 60% di foreste non gestite e 40% di foreste a gestione intensiva. Questa combinazione sarebbe in grado di supportare fino al 94% di tutte le specie campionate, come riportato dagli autori. È un esito a prima vista sconcertante, anzitutto perché comprende il 40% di gestione intensiva, che di solito associamo a una eccessiva semplificazione o degrado della biodiversità. La chiave è la complementarità: alcune specie e gruppi traggono vantaggio da habitat più aperti e fasi pionere della successione, che la gestione intensiva è in grado di mantenere (tagli recenti, più luce, microclimi diversi), mentre altri dipendono da grandi alberi, legno morto, microhabitat: elementi più frequenti nei boschi del tutto non gestiti.
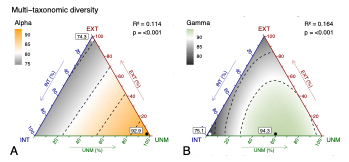
La parte che “pizzica” il dibattito europeo è anche un’altra: la gestione estensiva, nel dataset e per questo tipo forestale, non contribuisce alla conservazione della biodiversità quanto ci si aspetterebbe. Gli autori lo dicono esplicitamente: l’estensivo ha il contributo più debole nel complesso, e la diversità multi-tassonomica alfa (a scala di plot) risulta addirittura minima nei paesaggi “puramente estensivi” rispetto ai puramente intensivi o puramente non gestiti. È un risultato scomodo, perché molte strategie di selvicoltura naturalistica puntano proprio su quella fascia intermedia.
Scendendo nei dettagi, si vede che le risposte cambiano molto tra gruppi. I funghi lignicoli, per esempio, hanno massimi di diversità in paesaggi dominati dal non gestito, coerentemente con la dipendenza da legno morto e grandi alberi; per le piante vascolari, invece, l’intensivo promuove sia alfa sia gamma, e il minimo è nel non gestito, probabilmente perché una faggeta matura e chiusa lascia poca luce e poche nicchie per molte specie di sottobosco. Uccelli e coleotteri saproxilici mostrano benefici da combinazioni di non gestito e intensivo, con percentuali ottimali diverse tra loro, un’altra indicazione che “biodiversità” non è un unico numero.
Gli autori aggiungono un caveat importante: molte delle foreste “non gestite” europee sono tali da pochi anni o decenni, perché quasi tutte hanno una storia di gestione. Eppure, anche l’abbandono recente può già aumentare variabilità di diametri, presenza di alberi più grandi e volumi di legno morto rispetto alle gestite. Questo rende il risultato meno “utopico” del previsto sul breve periodo, ma lascia aperta la domanda di fondo che loro stessi pongono: ritirare dalla gestione il 60% del paesaggio è irrealistico con la domanda crescente di legno, la strada praticabile può diventare aumentare nei boschi gestiti in modo estensivo quelle caratteristiche strutturali che oggi mancano, cioè microhabitat, grandi alberi, legno morto, e la loro variabilità spaziale?

Questo chiude anche il filo dei cinque post: che si parli di carbonio, disturbi, composizione o pianificazione, la lezione ricorrente è che le foreste non si governano con un unico obiettivo e un’unica ricetta. Servono scelte esplicite su cosa si vuole massimizzare, su dove accettare i compromessi e su come misurare se la direzione è quella giusta, perché è in quella chiarezza, più che nelle parole d’ordine, che si gioca la credibilità della gestione forestale in un clima che cambia. E serve un filo diretto tra ricerca scientifica e gestione, che SISEF si impegnerà a tessere anche per il 2026.
Dipartimento di Scienze Agrarie e Ambientali - Produzione, Territorio, Agroenergia (DISAA)
Università degli Studi di Milano































